
забарвлені пластиди, які містять, в основному, червоні, помаранчеві й жовті пігменти. Вони, як правило, розвиваються з хлоропластів, мають приблизно такі самі розміри та форму, досить схожі й за структурою.
Усередині хромопласти заповнює строма, в якій іноді трапляються рудименти мембранної системи: окремі тилакоїди, скупчення трубочок. Однак замість системи фотосинтетичних мембран в них є структури, багаті на каротиноїди.
Хромопласти трапляються в пелюстках квіток деяких рослин (жовтеці, кульбаби та ін.), у плодах (шипшина, перець, горобина, помідори, кавун), рідше – у вегетативних органах (морква).
Фізіологічна функція хромопластів детально не вивчена. Яскраве забарвлення, можливо, принаджує комах для запилення та поширення насіння.
Лейкопласти – дрібні безбарвні пластиди, які переважно містяться в клітинах тканин, на які не потрапляє світло (сім'ядолі, ендосперм насіння, кореневища, бульби, корені), рідше – у тканинах на світлі (шкірка).
Основна функція лейкопластів – накопичення поживних речовин, які надходять у запасаючі органи.
Внутрішня мембрана лейкопластів формує поодинокі тилакоїди. Лейкопласти мають власну ДНК, рибосоми, а також різноманітні ферменти. Лейкопласти, в яких синтезується і відкладається крохмаль у незелених частинах рослин, називають амілопластами, рослинні олії – олеопластами, білки – протеїнопластами.
Амілопласти – найрозповсюдженіші лейкопласти, в них накопичується запасний крохмаль у вигляді зерен. Амілопласти є в запасних тканинах та органах, а саме: сім'ядолях, ендоспермі, бульбі, а також у кореневому чохлику.
Протеїнопласти містять у стромі запасний білок у вигляді кристалів або аморфної маси. Лейкопласти такого типу є рідкісним явищем (кореневі бульби орхідей).
Олеопласти трапляються ще рідше. У їхній стромі збираються жири (олії) у вигляді крапельок або пластоглобул (однодольні).
У кореневищах деяких видів півників у лейкопластах може накопичуватися крохмаль або жир.
Взаємоперетворення пластид. Усі пластиди – хлоропласти, хромопласти, лейкопласти – можуть розвиватися безпосередньо з пропластид, що містяться в клітинах твірної тканини.
Мітохондрії. По всій цитоплазмі клітин розсіяні численні мітохондрії, довжина яких – від одного до кількох мкм, а ширина – до 0,5 мкм. Мітохондрії – органоїди, присутні як у рослинних, так і у тваринних клітинах. У рослинній клітині може міститися від декількох десятків до декількох тисяч мітохондрій. Кількість їх залежить від віку і метаболічної активності клітин. Деякі водорості (хлорела, хламідомонада) мають у клітині лише одну велику мітохондрію.
Мітохондрії оточені подвійною ліпопротеїновою мембраною. Зовнішня мембрана вирізняється від внутрішньої співвідношенням білків і ліпідів, набором ферментів, проникністю. Внутрішня мітохондріальна мембрана утворює впинання – гребені, або кристи, розмір та кількість яких залежать від функціональної активності мітохондрій. Кристи утворюють велику поверхню для реакцій, що проходять на внутрішній мембрані. Простір між кристами заповнено основною речовиною – матриксом, у якому помітні гранулярний і фібрилярний компоненти. Гранули представлені рибосомами і фосфатом кальцію, фібрили – це нитки ДНК.
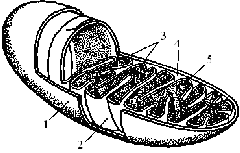
Рис. 1.5. Схема будови мітохондрії: 1 – зовнішня мембрана; 2 -внутрішня мембрана; 3 – кристи; 4 – міжмембранний простір; 5 – матрикс
Подібно до пластид мітохондрії є напівавтономними органоїдами, оскільки в них відбувається незалежний від ядра білковий синтез. У матриксі мітохондрій локалізована власна білоксинтезуюча система з кільцевих молекул ДНК, РНК і 70S рибосомами. Мітохондріальний геном кодує до 50 мембранних і матриксних білків.
Основна функція мітохондрій – енергетична. У них відбувається процес дихання – окиснення поживних речовин, насамперед вуглеводів, з виділенням енергії, що запасається у формі АТФ. Розклад вуглеводів починається в матриксі цитоплазми (гліколіз), а продовжується в матриксі мітохондрій (цикл Кребса). Синтез АТФ відбувається на внутрішній мітохондріальній мембрані.
Мітохондрії постійно рухаються й збираються там, де потрібна енергія. Тривалість життя мітохондрій – декілька діб. Мітохондрії розмножуються в клітинах шляхом поділу.
Детальне вивчення структури та геному мітохондрій дає можливість припустити, що вони, як і хлоропласти, походять від бактерій, які випадковим симбіозом прижилися в більших гетеротрофних клітинах – попередниках евкаріот.
Лізосоми – дрібні округлі органоїди діаметром 0,5-2,5 мкм. Вважають, що у тваринних клітинах їх більше, ніж у рослинних. Лізосоми оточені однією ліпопротеїновою мембраною, усередині знаходиться матрикс, що містить гідролітичні ферменти, які розщеплюють різні органічні речовини – білки, нуклеїнові кислоти, полісахариди та ін. Лізосоми утворюються з цистерн апарату Гольджі або з гладких мембран ЕПС. Функція лізосом – розщеплення речовин і окремих ділянок цитоплазми, тобто локальний автоліз.
У лізосомах розщеплюються «відпрацьовані» речовини й органоїди, що у даний момент онтогенезу вже виконали свої функції і не потрібні клітині. Продукти їх розпаду використовуються на побудову нових, необхідних їй речовин і структур.
У лізосомах розпадаються речовини, часто шкідливі, які потрапляють у клітину шляхом ендоцитозу. Продукти їх розщеплення використовуються клітиною.
При відмиранні клітин порушуються мембрани лізосом, їхні ферменти виходять у цитоплазму, відбувається автоліз усього живого вмісту, що спричиняє очищення порожнини мертвих клітин (трахеї і трахеїди ксилеми, клітини механічних тканин тощо).
Рибосоми – це дрібні клітинні органоїди, які можна побачити тільки під електронним мікроскопом. Рибосоми мають