
понад 600 родів. Поширені переважно у теплих морях і океанах. У прісних і солонуватих континентальних водоймах, а також в аерофітних умовах мешкає не більше 50 видів.
Морські багрянки – це переважно багатоклітинні великі рослини, зі складною будовою, які досягають у довжину від кількох сантиметрів до метра. Тіло має складну будову: розчленоване, нагадує розгалужені кущики або стрічковидні пластинки.
Серед морських багрянок є багато мікроскопічних форм, але лише найпримітивніші з них мають одноклітинні або колоніальні таломи. У високоорганізованих представників слань має складну тканинну будову. Периферійні шари слані складаються з дрібних корових клітин, багатих на хлоропласти. Внутрішні клітини більші за розміром, не мають хлоропластів і виконують різні функції.
Багатоклітинні таломи багрянок розгалужуються моноподіально, симподіально, дихотомічно. До субстрату водорості прикріплюються ризоїдами на нижньому боці талому, а також виростами-присосками або дископодібною підошвою.
Клітини червоних водоростей різноманітні за формою, з цілісною двошаровою оболонкою. Внутрішній шар оболонки складається з целюлози, зовнішній – з пектинових речовин. Серед пектинових речовин є колоїдні речовини – агар, карагінан, агароїди. Вони здатні розчинятися в киплячій воді з утворенням колоїдних розчинів. До групи пектинових речовин відносяться також колоїдні речовини, які містяться в клітинних оболонках і міжклітинниках багатьох морських багрянок (Phyllophora).
У морських високоорганізованих форм клітини багатоядерні, а у примітивних форм і репродуктивних клітин (спермаціїв, карпо- і тетраспор) одноядерні. Ядро маленьке, з ядерцем.
У цитоплазмі є один-два хлоропласти, у найпримітивніших форм з піреноїдом. Форма хлоропластів різна і не завжди постійна; вона може бути різною навіть в різних клітинах одного організму і змінюється з віком організму.
Червоні водорості мають складний набір пігментів. Поряд з хлорофілами а і d, a- і b-каротинами і кількома ксантофілами, хлоропласти червоних водоростей містять специфічні для даної групи рослин додаткові пігменти: червоний фікоеритрин і синій фікоціанін. Ці пігменти виявлені також у синьозелених і криптофітових водоростей. Різне кількісне співвідношення фікоеритрину і фікоціаніну визначає колір червоних водоростей. У прісноводних водоростей фікоеритрину мало або він відсутній, тому вони забарвлені переважно в сірий, темно-зелений, синьо-зелений або зелений колір.
Запасним полісахаридом є багрянковий крохмаль, який від йоду не синіє, а спочатку стає жовто-бурим, а потім червоним.
Більшість червоних водоростей – дводомні рослини, які мають статеві органи складної будови. Спорофіт і гаметофіт за будовою подібні. Розмножуються нестатевим і статевим шляхом.
Вегетативне розмноження одноклітинних і колоніальних форм відбувається поділом клітини. У багатоклітинних водоростей утворюються додаткові пагони, що перетворюються у нові рослини. Нестатеве розмноження у низькоорганізованих водоростей здійснюється моноспорами, тобто спорами, які утворюються по одній у клітині. Для більш високоорганізованих характерне нестатеве розмноження тетраспорами, які утворюються по чотири в клітині, що називається тетраспорангієм. І моноспори, і тетраспори без оболонки, без джгутиків. Потрапляючи у водне середовище, вони вкриваються оболонкою і проростають.
Моноспори і тетраспори проростають, як правило, в гаметофіти, на яких утворюються органи статевого розмноження, які мають складну будову.
Статевий процес оогамний. Лише одноклітинні і колоніальні багрянки статево не розмножуються. Гамети не мають джгутиків. Чоловічі гамети – спермації – утворюються в клітинах, які функціонують як чоловічі органи розмноження і називаються сперматангіями, або антеридіями. При заплідненні спермації викидаються назовні і пасивно переносяться течією води, а жіночі гамети – оогонії – залишаються на гаметофіті у жіночому статевому органі, який називається карпогоном і утворюється на кінцях бічних гілочок. Карпогон – це особлива клітина без хлоропласта. У бангієвих водоростей карпогон являє собою мало змінену вегетативну клітину, яка відрізняється від решти більшими розмірами і інтенсивнішим забарвленням. У флоридових нижня частина карпогону розширена, містить ядро, а верхня частина вузька, утворює трубчастий виріст – трихогіну, яка служить для вловлювання спермаціїв.
При заплідненні спермації прикріпляються до вершини трихогіни, оболонки їх в місці зіткнення розчиняються, і ядро спермація перетікає в порожнину трихогіни. Пересуваючись по ній, воно потрапляє в черевну частину карпогону і там зливається з його ядром. Після запліднення карпогон відділяється від трихогіни поперечною перегородкою, трихогіна відмирає.
Зигота без періоду спокою починає ділитися і утворює особливі спори – карпоспори, із яких розвиваються спорофіти (рис. 2.19).
У бангієвих після запліднення карпогон ділиться на 4-32 карпоспори, які у вигляді голих клітин виходять у воду. У флоридових у розвитку зиготи крім карпогону беруть участь інші спеціальні клітини.
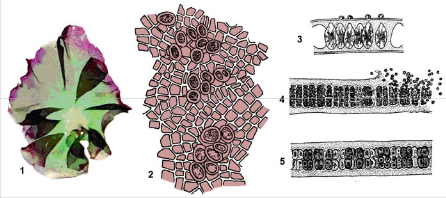
Рис. 2.19. Порфіра (Porphyra sp.) із пластинчастою структурою талому (1 – загальний вигляд; 2 – вигляд зверху, помітні карпоспори; 3 – 5 поперечний зріз пластинки: 3 – з карпогонами, 4 – з сперматангіями, 5 – з карпоспорами)
З диплоїдних карпоспор утворюється тетраспорофіт, який розмножується лише нестатево – тетраспорами. Тетраспори утворюються редукційним поділом. Вони гаплоїдні і при проростанні утворюють статеву генерацію – гаметофіт.
Таким чином, у більшості червоних водоростей розвиток проходить такі форми: гаметофіт, карпоспорофіт, тетраспорофіт. У цьому циклі є вільноживучі форми розвитку – тетраспорофіти і гаметофіти. Редукційний поділ відбувається при утворенні тетраспор, у зв'язку з чим тетраспори і гаметофіти, що утворюються з них, є гаплоїдною фазою, а зигота, карпоспори і тетраспорофіт – диплоїдною фазою.
Червоні водорості широко розповсюджені у всіх морях, але у напрямку високих широт загальна кількість видів багрянок поступово зменшується і різко падає в Арктичному