
поля – центральне, осьове і бічне.
Потовщення, що виступають над зовнішньою або внутрішньою поверхнею стулки називаються ребрами, вони забезпечують міцність панцира. У багатьох діатомових водоростей на зовнішній поверхні панцира утворюються виступи, щетинки, шипики, які збільшують його поверхню і служать для з'єднання клітин в колонію.
За будовою панцира діатомові розділяють на два класи: пенатні (Pennatophyceae) з двобічносиметричним панциром і центричні (Centrophyceae) з радіально-симетричним панцирем.
Велике біологічне значення має слиз, що виділяється діатомовими водоростями, який не лише сприяє утворенню колоній і прикріпленню водоростей до субстрату, але і захищає їх від висихання та дії інших несприятливих умов середовища.
Багато пенатних водоростей мають шов, який складається з однієї-двох щілин, що пронизують оболонку стулки від кінців до центра. Шов забезпечує взаємодію протопласту із зовнішнім середовищем і здатність до руху. Наявність шва – це прогресивна ознака, характерна для еволюційно молодих видів.
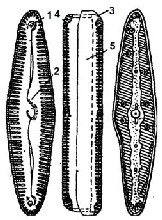
Рис. 2.14. Пенатна водорість пінулярія (Pinnularia sp.): 1 – вузол, 2 – шов, 3 – епітека, 4 – гіпотека, 5 – пояски
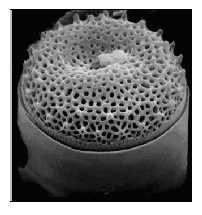
Рис. 2.15. Ореоли на панцирі таласіозіри (Thalassiosira sp.)
Діатомові мають виключно кокоїдний тип структури тіла. Розмір клітин від 4 до 2000 мкм. У відділі, за даними різних авторів, налічується 12-25 тис. видів.
Клітини живуть поодинці або об'єднані в колонії різного типу (рис. 2.16).
Ядро розміщене в центрі клітини в цитоплазматичному містку чи в пристінному шарі цитоплазми.
Хлоропласти діатомових водоростей різноманітні за формою, розмірами і положенням у клітині. У більшості діатомових хлоропласти дрібні, численні, у вигляді зерен або дисків. Лише у високоорганізованих пенатних хлоропласти великі, один чи два в клітині.
Хлоропласти забарвлені в жовто-бурий колір з різними відтінками. Забарвлення зумовлено пігментами: хлорофілами а і с, β-каротином, ε-каротином, рядом ксантинів. Пігментна система діатомових водоростей забезпечує фотосинтез на глибині до 50 м завдяки додатковим пігментам з групи фукоксантинів, які передають поглинену ними енергію хлорофілу а. Після загибелі клітини бурі пігменти розчиняються у воді і зелений хлорофіл стає яскраво помітним. У гетеротрофних безбарвних діатомових водоростей (Nitzschia) пігменти відсутні.
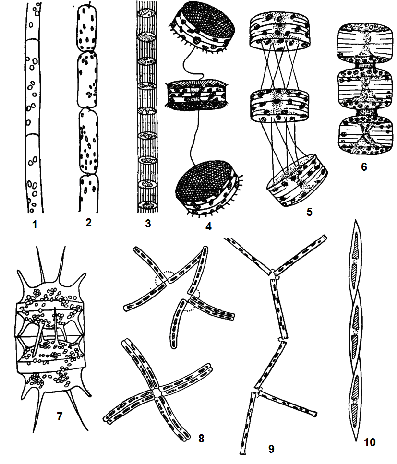
Рис. 2.16. Типи колоній діатомових: 1 – ниткоподібна колонія (Leptocylindrus danicus); 2-3 – ланцюжкові колонії (Rhizosolenia fragilissima, Sceletonema costatum); 4 – гнучкий ланцюжок (Thalassiosira excentrica); 5 – гнучкий ланцюжок (Thalassiosira polychorda); 6 – прямий ланцюжок (Porosira glaciatus); 7 – прямий ланцюжок, утворений за допомогою виростів (Biddulphia mobiliensis); 8 – зигзагоподібний ланцюжок (Thalassionema nitzchioides); 9 – зигзагоподібний ланцюжок (Diatoma elongatum); 10 – нитковидна колонія (Nitzschia seriata)
У периферичній частині клітини знаходиться силікалема – складна система цистерн і трубочок, у яких виявлені елементи кремнієвого панцирю. Зовнішня мембрана цистерн силікалеми тісно прилягає до плазмалеми. Функції силікалеми – формування панцирю і утворення нової плазмалеми.
Основний спосіб розмноження діатомових водоростей – вегетативний поділ клітини надвоє. Найінтенсивніше розмноження діатомових спостерігається навесні і восени. Перед поділом клітини у діатомей починають нагромаджувати в протопласті краплинки олії, завдяки чому він значно збільшується в об'ємі, епітека і гіпотека панцира розходяться. Протопласт ділиться на дві рівні частини. Ядро ділиться мітозом, утворюючи два дочірніх.
Після закінчення поділу кожна з дочірніх клітин одержує від материнської лише одну стулку панцира і добудовує обов'язково внутрішню, тобто гіпотеку. Після багаторазових поділів спостерігається поступове зменшення розмірів клітин водоростей. У деяких видів вони зменшуються в 3 рази у порівнянні з первинними. Відновлення первинних розмірів клітин відбувається під час проростання спор і в результаті статевого процесу.
Перед початком статевого процесу дві клітини зближуються, покриваються загальним слизистим чохлом і розсовують стулки. Після запліднення утворюється зигота, яка без стадії спокою починає рости, збільшується в розмірах і перетворюється на ауксоспору («спору, що росте»). Клітина, що утворюється з ауксоспори, називається ініціальною і більша за вихідну клітину.
У діатомових виявлено декілька типів статевого процессу: ізогамний, анізогамний (гетерогамний), оогамний, автогамний.
Всі діатомові водорості диплоїдні організми, а гаплоїдна фаза ядра у них спостерігається тільки перед злиттям ядер у ауксоспорі.
У несприятливих умовах діатомові водорості переходять у стан спокою. Тоді протопласт пересувається до одного з кінців клітини і, унаслідок втрати клітинного соку, сильно стискається. Життєдіяльність цих клітин поновлюється при настанні сприятливих умов. Деякі прісноводні планктонні види в зимовий період занурюються на дно водойми, де перебувають в стані спокою або зниженої життєдіяльності до початку нового вегетаційного періоду.
Діатомові водорості широко розповсюджені і населяють різні біотопи: прісні і солоні, стоячі і проточні водойми, вологі скелі, ґрунт і навіть орні землі.
У водних екосистемах діатомові – цілорічні домінанти. У відкритій частині океану діатомові можуть розповсюджуватися до глибини 80-350 м, де утворюють глибоководний «тіньовий планктон». Це обумовлено здатністю деяких видів переходити на гетеротрофне живлення при великій кількості органічних речовин у воді. У планктоні прісних водойм переважають пенатні діатомові; глибина їх розповсюдження залежить від прозорості води.
У бентосних фітоценозах теж переважають пенатні (Navicula, Campylodiscus, Pinnularia, Gyrosigma). Переважно вони мешкають на глибині не більше 50 м.
Діатомові водорості – основна ланка трофічних ланцюгів водних екосистем. Вони відіграють основну роль