
їх коливається від 1-2 до багатьох. Деякі види роду евглена (Euglena), виробляють велику кількість червоного пігменту (астаксантину), що відіграє певну роль у регулюванні процесу засвоєння світла хлоропластами.
Основною запасною речовиною евгленових водоростей є полісахарид парамілон, який концентрується навколо піреноїда або утворюється безпосередньо в протоплазмі.
Вічко, або стигма, розташоване на передньому кінці клітини поблизу джгутикового апарату (рис. 2.8).
Ядро велике, нуклеоплазма містить одне або декілька ядерць.
Живлення у евгленових автотрофне, мезотрофне, гетеротрофне, що обумовлює наявність значного числа безбарвних форм.
Фототрофний тип живлення характерний для більшої частини представників порядку евгленових: евглен (Euglena), трахеломонасів (Trachelomonas), стромбомонасів (Strombo-monas), лепоцинклісів (Lepocinclis) і інших зелених форм. Сапрофітний тип живлення переважає у безбарвних представників цього ж порядку: астазій (Astasia), дистигм (Distigma), меноїдіумів (Menoidium) і інших форм, що не мають хлоропластів.
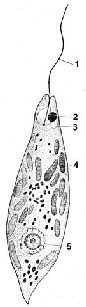
Рис. 2.8. Евглена зелена (Euglena viridis): 1 – джгутик; 2 – вічко (стигма); 3 – скоротлива вакуоля; 4 – хроматофори; 5 – ядро
Для родів перанема (Peranema), гетеронема (Heteronema), ентосифон (Entosiphon) і інших, що об'єднуються в порядок перанемових, характерний голозойний (анімальний) тип живлення: заковтування оформлених частинок органічної речовини або інших дрібніших організмів.
Розмноження відбувається шляхом поздовжнього поділу особини на дві.
Евгленові – це переважно прісноводні водорості, досить широко розповсюджені у водоймах з повільною течією. Більшість з них розвивається в поверхневих або придонних шарах води. Деякі види повзають по дну. Як виняток серед евгленових водоростей евтрепція (Eutreptia), яка віддає перевагу морським або солонуватим водоймам. Серед евгленових виявлені і паразитичні форми, що живуть в кишечниках олігохет, нематод, копепод, амфібій, на зябрах риб.
Господарське значення евгленових полягає у здатності деяких з них до міксотрофного живлення, що забезпечує їх участь у самоочищенні вод.
Ряд видів використовуються як біологічні індикатори забруднення водойм, у медичній практиці як модельні об'єкти для з'ясування дії різних лікувальних препаратів.
Багато видів евглен (Euglena) володіють здатністю до масового розвитку, викликаючи «цвітіння» континентальних вод.
2. Динофітові водорості (Dinophyta)
Динофітові водорості, або динофлагеляти, – це переважно одноклітинні водорості з монадним типом структури. Проте є і колоніальні і, зрідка, нитчасті форми. Клітини динофітових двобічно симетричні або асиметричні, переважно округлої, еліпсоїдної, яйцевидної, рідше грушовидної або іншої форми тіла. Деякі види мають форму тіла у вигляді півмісяця або піраміди. Клітини зазвичай забарвлені в темно-бурий, червоний, іноді в жовтий або жовто-зелений колір, є також безбарвні форми.
Розміри коливаються в межах від 6 до 1500 мкм. Серед прісноводних представників найбільших розмірів досягають види роду цераціум (Ceratium) – до 450 мкм. Відомо близько 300 видів динофітових, що мешкають у прісних і морських водах.
Клітинний покрив у найбільш примітивних форм представлений гладким тонким і ніжним перипластом. Переважна більшість динофітових вкриті целюлозною оболонкою, яка складається з пластинок (щитків), які часто утворюють панцир. Він складається з трьох основних частин: верхньої – епітеки, нижньої – гіпотеки і середньої – пояска, що сполучає обидві частини в суцільний панцир (рис. 2.9.). Епітека і гіпотека складаються з пластинок і щитків, сполучених у певному порядку швами. Число і розміщення пластинок, а також наявність шипиків на поверхні належать до важливих систематичних ознак.

Рис. 2.9. Цераціум (Ceratium): 1 – поясок, 2 – апікальний виріст, 3 – задній джгутик, 4 – антапікальний виріст
Монадні динофітові мають по два джгутика різної довжини і будови – поперечний і подовжній. Поперечний джгутик має вид хвилястої нитки, виходить з верхньої джгутикової пори і опоясує клітину вліво. Цей джгутик розташований в поперечній борозенці. Подовжній – рульовий – джгутик у вигляді тонкої нитки виходить з нижньої джгутикової пори, і у більшості видів розташований в подовжній борозенці панцира. Він тягнеться уздовж клітини і забезпечує поступальний рух.
Динофітовим властива своєрідна будова ядра, яка називається динокаріоном, в якій хромосоми кріпляться до ядерної мембрани. Хромосоми, на відміну від інших еукаріот, без гістонів і складаються з мікрофібрил, що містять чисту ДНК. Під час мітозу ядерна оболонка не зникає і веретено поділу не утворюється. Це ядро мезокаріотного типу, яке вважається проміжним між нуклеоїдом прокаріотів і справжнім ядром еукаріотів.
Розмноження у динофітових водоростей здійснюється в основному поділом клітин в рухомому стані або зооспорами і апланоспорами. У деяких видів описаний статевий процес, що відбувається шляхом злиття дорослих клітин або джгутикових клітин і апланоспор. За несприятливих умов у багатьох панцирних видів утворюються товстостінні цисти (Ceratium согnutum).
Через нез’ясоване таксономічне положення в системі органічного світу, володіючи ознаками як рослин, так і тварин, динофітові привертають увагу ботаніків і зоологів. Цю таксономічну групу розглядають або у складі фітопланктону, або як компонент протозоопланктону. Одні і ті ж види (при збереженні їх видових і родових назв) входять до складу двох систем – ботанічної і зоологічної, підкоряючись двом різним номенклатурним кодексам.
Динофітові – це одна з домінуючих груп мікроводоростей у водних екосистемах. Це типові мешканці пелагіалі морів і океанів, хоча зустрічаються і в прісних континентальних водоймах.
Динофітові мають високу різноманітність у морських екосистемах