
які утримують гілочки у вертикальному положенні. Усі бурі водорості прикріплюються до грунту чи інших водних рослин за допомогою ризоїдів, базальних дисків.
У високоорганізованих бурих водоростей (Laminaria, Fucus, Sargassum) таломи диференційовані і нагадують квіткові рослини. У них є стебло-, листко- і коренеподібна частини, деякі представники мають повітряні міхурі, що утримують гілки у вертикальному положенні. Слань має примітивні тканини: асиміляційну, запасаючу, механічну і провідну.
Галуження бурих водоростей моноподіальне. Бічні гілочки чергові розсіяні або супротивні. При швидкому доростанні їх до розмірів головної нитки виникає дихотомоподібне галуження.
Клітини бурих водоростей одноядерні. Оболонка їх складається з внутрішнього целюлозного шару і зовнішнього пектинового. Целюлоза бурих водоростей за своїми властивостями відрізняється від целюлози квіткових рослин, і тому її іноді називають альгульозою.
Сусідні клітини сполучаються за допомогою пop і плазмодесм.
Хлоропластів декілька, вони маленькі, дископодібні, стрічкоподібні, пластинчасті з дуже дрібними піреноїдами. До їх складу входять пігменти: хлорофіли а і с, каротини і ксантофіли, особливо фукоксантин інтенсивно бурого кольору. Різні співвідношення цих пігментів обумовлюють забарвлення бурих водоростей від оливково-жовтуватого до темно-бурого, майже чорного кольору.
Клітини бурих водоростей мають одну велику або декілька дрібних вакуолей. Крім того, зустрічаються фізоди – це дуже дрібні вакуолі (до 4 мкм в діаметрі) із таніноподібною речовиною.
Запасною поживною речовиною є полісахарид ламінарин, шестиатомний спирт маніт і в невеликих кількостях олія.
Розмножуються бурі водорості вегетативно, нестатевим і статевим способами.
Вегетативне розмноження відбувається при випадковому відокремленні гілок від слані. Спеціальні бруньки вегетативного розмноження є лише у видів роду сфацелярія (Sphacelaria).
Нестатеве розмноження здійснюється зооспорами, нерухливими тетраспорами і моноспорами. Зооспори і гамети утворюються у одно- і багатогніздих вмістищах (рис. 2.24).
Статевий процес ізо-, гетеро- і оогамний. Гамети зазвичай утворюються в багатогніздних гаметангіях, по одній в кожній камері.
Більшість бурих водоростей мають самостійні форми розвитку – спорофіт і гаметофіт або гаметоспорофіт (рис. 2.25). Гаметофіт і спорофіт можуть мати як однакову будову і розміри, так і різну, тобто у бурих водоростей існує ізоморфна і гетероморфна зміна форм розвитку. Найчастіше більші розміри і складнішу будову має спорофіт.
До бурих водоростей, що не мають зміни форм розвитку, а лише зміну ядерних фаз, відносяться представники родин фукусових (Fucaceae), цистозейрових (Cystoseiraceae) і саргасових (Sargassaceae).
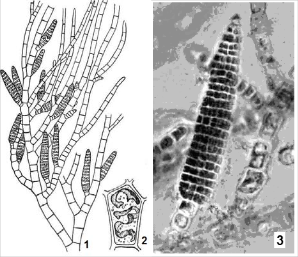
Рис. 2.24. Ectocarpus siliculosus: 1 – фрагмент талому з багатогніздними спорангіями; 2 – клітина із стрічкоподібними хлоропластами; 3 – спорангій (мікрофотографія)
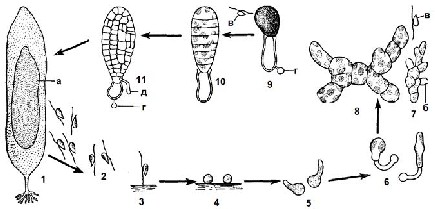
Рис. 2.25. Цикл розвитку ламінарієвих водоростей: 1 – талом зі спорами: а – пляма спорангіїв; 2 – плаваючі зооспори; 3 – зооспора прикріпилася переднім джгутиком до грунту; 4 – ембріоспори; 5 – ембріоспори з проростковою трубкою; 6 – утворення перших клітин гаметофітів (зліва – жіночий, справа – чоловічий); 7 – чоловічий гаметофіт: б – антеридій, в – антерозоїди; 8 – багатоклітинний жіночий гаметофіт; 9 – одноклітинний жіночий гаметофіт після виходу яйцеклітини з оогонію: г – оболонка ембріоспори; 10-11 – початок розвитку слані спорофіта: д – первинний ризоїд
Нормальне відтворення водоростей можливе лише статевим шляхом. Статевий процес – типова оогамія. Статеві органи розвиваються в концептакулах. Із стінки концептакула виростають довгі волоски – парафізи, що заповнюють майже всю його порожнину. Особливо довгі волоски розвиваються в жіночих концептакулах, де вони у вигляді пучка видаються з отвору концептакула. Серед цих волосків розвиваються оогонії і антеридії.
Майже усі бурі водорості винятково морські рослини. Найбільші за розмірами їх форми (ламінарієві, деякі фукусові) зустрічаються у морях приполярної і полярної зон. Тут для них сприятливі помірна і низька температура води і підвищений у порівнянні з водами тропічних морів вміст біогенних речовин. Селяться вони переважно на скелях і камінні на глибині 6-15 м.
Серед бурих водоростей є види з ефемерними, однорічними і багаторічними таломами. На тривалість існування таломів великий вплив мають екологічні умови. Багаторічні таломи бурих водоростей бувають декількох типів. У одних водоростей талом багаторічний, щороку відмирають тільки пагони, на яких розвивалися органи розмноження (Fucales), у інших (Laminariales) багаторічними є стовбур і органи прикріплення, пластинчаста частина – однорічна.
У деяких тропічних видів саргасових водоростей багаторічним буває лише диск, що служить для прикріплення талому.
У розвитку бурих водоростей спостерігається сезонність. Так, у помірних субарктичних поясах вони найбільше відростають влітку, хоч посилене збільшення їх сланей починається вже наприкінці зими, коли температура води близько 0°С. У тропічних морях масовий розвиток бурих водоростей спостерігається взимку.
Найскладніша будова у представників порядку ламінарієвих.
Роль у природі бурих водоростей надзвичайно велика. Вони є одним з основних джерел органічної речовини в прибережній зоні, особливо в морях помірних і приполярних широт, де їх біомаса може досягати десятків кілограмів на 1 м2. Крім того, зарості бурих водоростей служать місцем розмноження, укриття і живлення багатьох прибережних тварин; вони створюють також умови для поселення мікроскопічних і макроскопічних водоростей інших систематичних груп.
Велике і господарське значення бурих водоростей, особливо як сировина для отримання різних речовин (наприклад, альгінатів – солей альгінової кислоти, зокрема альгінату натрію).