
тому що розчинені у воді кисень і вуглекислий газ стебла поглинають усією поверхнею.
Часто замість замикаючих клітин продиху видно їх ініціальну клітину. Клітини епідерми містять хлоропласти і здійснюють фотосинтез, використовуючи промені світла, що проникають крізь шар води. Первинна кора займає велику частину діаметра стебла. Паренхіма кори пухка, з великими міжклітинниками, нерідко перетворюється в аеренхіму. Часто повітроносні порожнини розділяються перегородками з дрібних клітин із хлоропластами, а довгі міжклітинники – діафрагмами, у яких між клітинами є невеликі повітряні порожнини. Вони пропускають повітря, але перешкоджають проходженню води, що у випадку ушкодження тканин стебла не дозволяє воді заповнити повітряні порожнини.
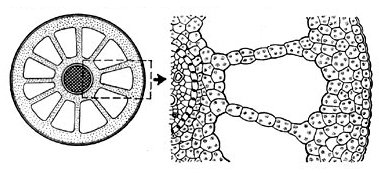
Рис. 1.6. Будова стебла водопериці: праворуч – при великому збільшенні; аеренхіма – видно великі повітроносні порожнини
Діаметр центрального циліндра у водних рослин невеликий. Провідні пучки сильно зближені й у більшості водних рослин перетворилися на один пучок, що складається з тяжа ксилеми і тяжа флоеми. Причому ксилема зазвичай розвинута слабко або зовсім відсутня, і на її місці утворюється повітряна порожнина. Це пов'язано з тим, що стебло поглинає воду всією поверхнею і не треба проводити її по стеблу. Механічної тканини в таких стеблах мало або вона зовсім відсутня. Рослину підтримує вода, тим більше, що велика кількість повітроносних порожнин надає їй плавучості. Камбій у стеблах, занурених у воду, працює дуже слабко, і вторинна будова, як правило, не формується.
6. Форми коренів і типи кореневих систем
За походженням розрізняють головний, бічні й додаткові корені.
Головний корінь (вісь першого порядку) утворюється тільки із зародкового корінця насінини.
Додаткові корені беруть початок від зони кущіння зародка, від стебла і листків або їхніх видозмін. Від головного й додаткових коренів відходять бічні корені – осі другого й наступних порядків галуження. При вирощуванні рослин для збільшення маси додаткових коренів у поверхневих родючих шарах ґрунту на приземні ділянки стебел нагортають купи землі. Щоб збільшити масу бічних коренів, проводять пікірування проростків (переважно при вирощуванні культурних рослин розсадою).
Головний, бічні й додаткові корені разом становлять кореневу систему. Розрізняють стрижневу, мичкуватута змішану кореневі системи. Стрижневу кореневу систему має більшість представників двосім'ядольних рослин (люцерна, соя, горох, соняшник тощо).
Мичкувата коренева система характерна для рослин класу односім'ядольних (жито, пшениця, овес, кукурудза та ін.). У цих рослин головного кореня немає (а якщо він зберігся, то за розмірами не вирізняється серед додаткових), і система представлена мичкою додаткових коренів, що утворилися від вузла кущіння. Змішана коренева система характерна здебільшого для представників трав'янистих двосім'ядольних рослин (суниці, полуниці), в яких відносно добре розвинені усі три типи коренів.
За формою корені дуже різноманітні: ниткоподібні, конусоподібні, веретеноподібні, бульбоподібні тощо.
Корені рослин проникають у ґрунт на велику глибину, досягаючи ґрунтових вод. Наприклад, головний корінь верблюжої колючки (рослина посушливих напівпустель з родини бобових) досягає глибини 20 м, головний корінь дворічного дуба – 2 м. На глибину 2 м проникають кореневі системи злакових рослин, а трав'янистих бобових – на 10 м і більше. Загальна довжина всіх коренів дуже велика. Коренева система рослин значно краще розвинена, ніж надземна частина. Наприклад, корені будяку заглиблюються більш як на 6 м за висоти надземної частини 1 м. За звичайних умов вирощування хлібні злаки (пшениця, овес, ячмінь) мають кореневу систему загальною довжиною 20 км, гарбузи – близько 25 км, а загальна довжина кореневої системи жита, яке вирощували в дослідних умовах, сягала 63,2 км. Діаметр кореневої системи плодових дерев у 2-5 разів більший за діаметр крони.
Корінь наростає своєю верхівкою. При обриві кореня спостерігається могутній розвиток бічного коріння. Ця особливість кореня використовується при пікіруванні. При пікіруванні пересаджують рослину і вкорочують головний корінь на 1/2-1/3 його довжини, що викликає рясне розгалуження частини, що залишилася, забезпечуючи розростання кореневої системи.
7. Анатомічна будова кореня
На поздовжньому зрізі молодого кореня можна бачити декілька зон, які добре виявляються.
Зона поділу клітин знаходиться на самому кінчику кореня. Вона складається з меристематичної тканини і являє собою конус наростання кореня. Розміри її дуже малі – до 1 мм.
Зона поділу вкрита кореневим чохликом, який захищає ніжні меристематичні клітини від ушкодження твердими частинками ґрунту.
Кореневий чохлик являє собою ковпачок із паренхімних клітин. Він утворюється зовнішнім шаром клітин апікальної меристеми. Тривалість життя його клітин 2-9 діб, залежно від фізичного стану ґрунту і виду рослин. Окрім захисту меристематичних клітин кореневий чохлик сприяє проникненню кореня в глибину. Він виділяє слиз, яким обволікається корінь, що полегшує його просування між частинками ґрунту, а також забезпечує реакцію кореня на гравітацію (позитивний геотропізм), яка визначає напрямок росту кореня.
Кореневий чохлик є у всіх сухопутних рослин. У водних рослин при рості їх коріння у воді чохлик не утворюється. При вкоріненні їх в ґрунті на кінці коріння розвивається чохлик.
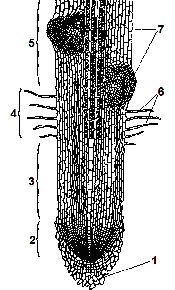
Рис. 1.7. Зони кореня: 1 – кореневий чохлик; 2 –